°ЧҪйЛШ-12ЈЁIL-12Ј©өД·ЦЧУҪYҳӢЎўЧчУГј°Х{№қҷCЦЖ
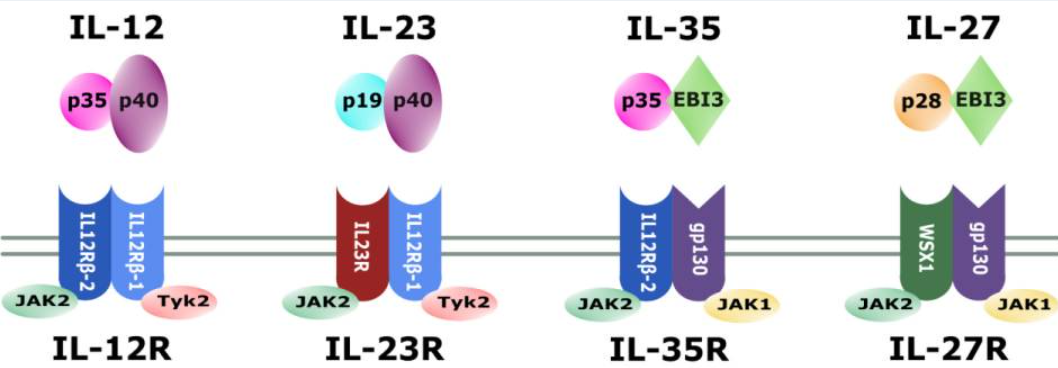
IL-12КЗТ»·N®җФҙ¶юҫЫуwјҡ°ыТтЧУЈ¬УЙαжңp35ЈЁјs35 kDaЈ©әНβжңp40ЈЁјs40 kDaЈ©ҪMіЙЎЈЯ@ғЙӮҖҒҶ»щ·Ц„eУЙІ»Н¬»щТтҫҺҙaЈЁИЛоҗ»щТтО»УЪөЪ3М–әН5М–ИҫЙ«уwЈ©ЎЈЦөөГЧўТвөДКЗЈ¬p35ұҫЙнІ»ҫЯӮдӘҡБў·ЦГЪөДДЬБҰЈ¬ұШнҡЕcp40ҪYәПәуІЕДЬХэҙ_ХЫҜBІў·ЦГЪһйЙъОп»оРФөДIL-12®җЩ|¶юҫЫуwЈЁНЁіЈ·QһйIL-12p70Ј©ЎЈIL-12јТЧеЖдЛьіЙҶTТІІЙУГоҗЛЖҪYҳӢЈәАэИзIL-12p40ЕcБнТ»αжңIL-23p19ҝЙҪMіЙIL-23Ј¬¶шp35ТІҝЙЕcEBI3ЈЁEBІЎ¶ҫХTҢ§»щТт3Ј©ҪYәПРОіЙГвТЯТЦЦЖРФјҡ°ыТтЧУIL-35ЎЈЯ@·N№ІПнҒҶ»щҷCЦЖК№өГIL-12ЕcIL-23өИјҡ°ыТтЧУФЪҪYҳӢЙПҫЯУРУHҫүкPПөЎЈ
IL-12p40ҒҶ»щФЪСӘЗеәНјҡ°ыЕарBЙПЗеЦРіЈТФЯ^БҝҶОуwРОКҪіц¬FЈ¬ІўДЬРОіЙp40-p40Н¬¶юҫЫуwЎЈСРҫҝұнГчЈ¬p40Н¬¶юҫЫуwҝЙЮЧҝ№IL-12p70ҙЩЯMIFN-γ®aЙъөД»оРФЈ¬ұM№ЬЖдЙъАнТвБxЙРОҙНкИ«ГчБЛЎЈp40ҶОуwЯ^Бҝ·ЦГЪҝЙДЬ…ўЕcХ{№қГвТЯЖҪәвЈ¬¶шp35өДұнЯ_„tёьјУҸV·әЈ¬ө«ғHФЪЕcp40ҪYәПәуІЕДЬ°l“]ЧчУГЎЈ
IL-12КЬуwөДҪMіЙЕcұнЯ_·ЦІј
IL-12өДјҡ°ыұнГжКЬуwУЙғЙӮҖҝзДӨҒҶ»щҳӢіЙЈәIL-12Rβ1әНIL-12Rβ2ЎЈЖдЦРIL-12Rβ1ЕcIL-23КЬуw№ІУГЈЁIL-23РЕМ–ҸНәПуwУЙIL-12Rβ1әНҢЈҢЩөДIL-23RҳӢіЙЈ©Ј¬¶шIL-12Rβ2„tМШ®җРФөШЕcIL-12ҪYәПРОіЙёЯУHәНБҰКЬуwЎЈIL-12Rβ1ЕcIL-12Rβ2Н¬ҢЩУЪgp130јТЧеЈ¬Жд°ыНвҪYҳӢУтЕcgp130ө°°ЧҫЯУРН¬ФҙРФЎЈФЪГвТЯјҡ°ыЦРЈ¬ғЙ·NҒҶ»щЦчТӘұнЯ_ФЪ»о»ҜөДTБЬ°Нјҡ°ыәНNKјҡ°ыұнГжЎЈ»о»Ҝ о‘BПВөДBјҡ°ыТІҝЙұнЯ_IL-12КЬуwЈәФЪIFN-γәНIL-12№ІН¬ЧчУГПВЈ¬ИЛBјҡ°ыЙПХ{IL-12Rβ2ұнЯ_Ј¬ІўФЪIL-12ҙМјӨПВ°lЙъSTAT4БЧЛб»ҜЈ¬ұнГчBјҡ°ыФЪМШ¶Ё—lјюПВТІДЬн‘‘ӘIL-12РЕМ–ЎЈЦөөГТ»МбөДКЗЈ¬IL-12Rβ1өДУHәНБҰЭ^өНЈЁKd ~2–6 nMЈ©Ј¬Гҝјҡ°ыұнЯ_”өБҝЭ^¶аЈЁјs1000–5000ӮҖКЬуwЈ©Ј¬¶шIL-12Rβ2„tһйёЯУHәНБҰКЬуwЈЁKd ~5–20 pMЈ©Ј¬ұнЯ_”өБҝПаҢҰЭ^ЙЩЈЁјs100–1000ӮҖЈ©ЎЈ
ФЪIL-12РЕМ–ӮчҢ§ЦРЈ¬IL-12Rβ1әНIL-12Rβ2өД°ығИ…^·Ц„eҪYәПІ»Н¬өДJanusјӨГёЈәIL-12Rβ1ЦчТӘҪYәПTyk2Ј¬¶шIL-12Rβ2ҪYәПJAK2ЎЈIL-12ЕcКЬуwҪYәПәуЈ¬КЬуwПакPөДJAK2әНTYK2ұ»»о»ҜІў°lЙъЧФБЧЛб»ҜЈ¬ЯM¶шБЧЛб»ҜКЬуw°ығИ…^ЙПөДАТ°ұЛбЎЈIL-12Rβ2ЙПкPжIөДБЧЛб»ҜО»ьcЈЁИзY800Ј©һйSTAT4МṩҪYәПО»ьcЈ¬РОіЙРЕМ–ЖҪЕ_ЎЈҙЛНвЈ¬STAT4ҝЙұ»јӨГёјТЧеЦРөДp38 MAPKБЧЛб»ҜЈЁҪz°ұЛбО»ьcЈ©Ј¬Я@Т»РЮп—ҢҰSTAT4өДЮDдӣ»оРФәНIFN-γ®aЙъЦБкPЦШТӘЎЈҙЛНвЈ¬IL-12ЯҖҝЙјӨ»оPI3K/AktНЁВ·Ј¬ҙЩЯMTјҡ°ыФцЦіЎЈ
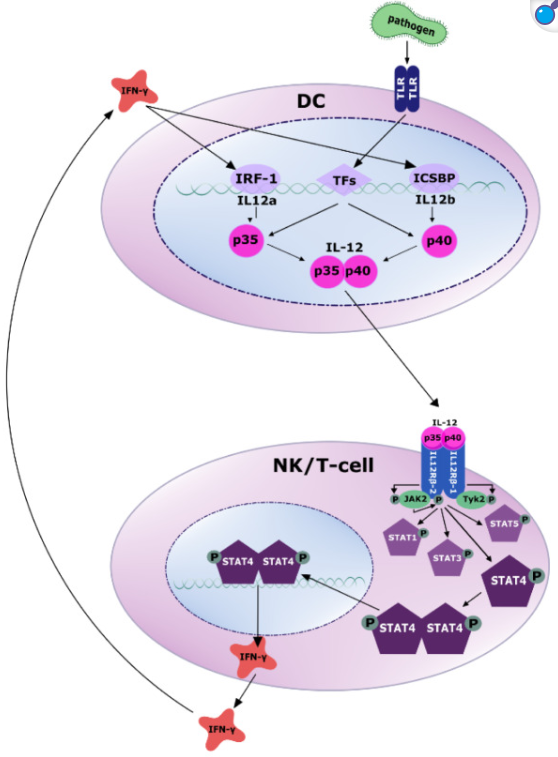
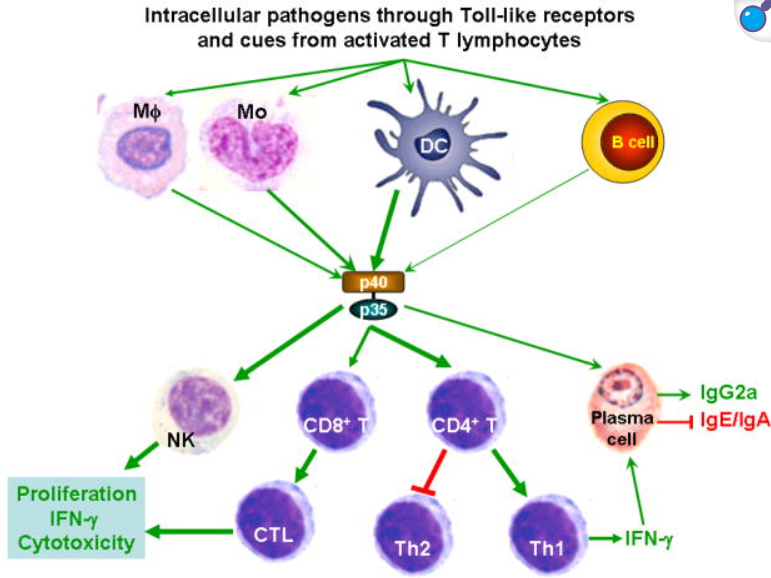
ҝ№ФӯіКЯfјҡ°ы (APC)Ј¬АэИзҳдН» ојҡ°ыЈ¬НЁЯ^ Toll ҳУКЬуwЈЁTLR) ёРЦӘ PAMPЈЁІЎФӯуwПакP·ЦЧУДЈКҪЈ©ЎЈлSә󣬶а·NЮDдӣТтЧУұ»јӨ»оЈ¬ХTҢ§ IL-12p35 әН IL-12p40 өДЮDдӣЈЁёь¶аФ”ЗйХҲ…ўйҶФӯОДЈ©ЎЈ·ЦГЪөД IL-12 ®җ¶юҫЫуwЕc NK јҡ°ыәН T јҡ°ыЙПөДКЬуwҪYәПЈ¬ДјјҜАТ°ұЛбјӨГё JAK2 әН TYK2Ј¬ІўНЁЯ^АТ°ұЛбБЧЛб»ҜјӨ»о JAK2ЎЈ»о»ҜөД JAK2 БЧЛб»Ҝ IL12Rβ2 ҒҶ»щЈ¬ЯM¶шНЁЯ^БЧЛб»ҜјӨ»о STAT4ЎЈлSәуЈ¬БЧЛб»ҜөД STAT4 °lЙъН¬¶юҫЫ»Ҝ»т®җ¶юҫЫ»ҜЈ¬ЮDО»ЦБјҡ°ыәЛЈ¬ІўНЁЯ^ҪYәП°Р DNA Х{ҝШ»щТтЮDдӣЎЈЦчТӘ°Р»щТтКЗ IFN-γЈ¬Жд·ҙЯ^ҒнНЁЯ^ IRF-1 әН ICSBP ХTҢ§ IL-12 ®aЙъөДЮDдӣјӨ»оЎЈ
IL-12РЕМ–өДШ“·ҙрҒХ{ҝШһй·АЦ№Я^¶ИөДСЧ°Y·ҙ‘ӘЈ¬IL-12РЕМ–НЁВ·КЬөҪ¶а·NШ“·ҙрҒҷCЦЖөДЦЖјsЎЈТЦЦЖРФјҡ°ыТтЧУРЕМ–·ЦЧУSOCS-3КЗIL-12Ш“Х{№қөДкPжI·ЦЧУЦ®Т»ЎЈSOCS-3өДSH2ҪYҳӢУтДЬүтЧR„eІўҪYәПБЧЛб»ҜөДIL-12Rβ2ЙПөДY800О»ьcЈ¬ҸД¶шёӮ ҺРФЧи”аSTAT4ЕcКЬуwөДҪYәПЎЈСРҫҝп@КҫЈ¬ФЪёЯұнЯ_SOCS-3өДTh2јҡ°ыЦРЈ¬IL-12ҪйҢ§өДSTAT4»о»Ҝұ»ҸҠБТТЦЦЖЈ¬ұнГчSOCS-3ФЪTh2јҡ°ыЦР·АЦ№IL-12РЕМ–ҪйҢ§өДTh1ҳO»ҜЎЈіэSOCS-3НвЈ¬ЖдЛыSOCSө°°Ч»тө°°ЧАТ°ұЛбБЧЛбГёТІҝЙДЬ…ўЕcIL-12РЕМ–өДХ{ҝШЈ¬ө«SOCS-3ҢҰIL-12Rβ2өДТЦЦЖЧчУГКЗТСЦӘөДҪӣөдҷCЦЖЎЈҙЛНвЈ¬IL-12ДЬХTҢ§өДIFN-γәНЖдЛьПВУОР§‘ӘТІҝЙйgҪУ·ҙрҒХ{№қIL-12өДЙъіЙЈ¬ө«ЦчТӘөДјҡ°ығИШ“·ҙрҒИФҡw№ҰУЪSOCS-3өИ·ЦЧУөДЧчУГЎЈ
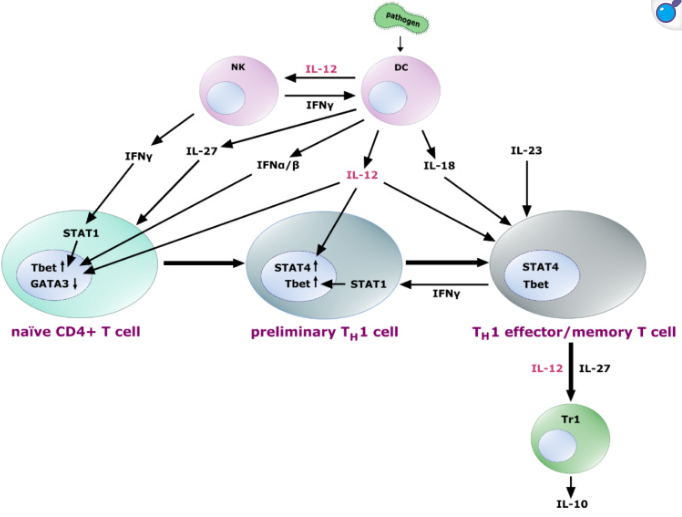
ұ©В¶УЪ IL-27 өДУЧЦЙ T јҡ°ыұнЯ_ IL-12 КЬуw®җ¶юҫЫуwЈ¬К№јҡ°ыҢҰ IL-12 өДУ°н‘ГфёРЎЈIL-12 ЕcҒнЧФ NK јҡ°ыәН·ҙрҒ»ШВ·өД IFN-γЈЁIFN-αәН IFN-βЈ©№ІН¬ХTҢ§ЮDдӣТтЧУ TbetөДЙПХ{әН GATA3өДПВХ{Ј¬ҸД¶шіхІҪҢҚ¬F TH1 ·Ц»ҜЎЈЯMТ»ІҪұ©В¶УЪ IL-12 •юҢ§ЦВЯ@Р©ФзЖЪ TH1 јҡ°ыЦР STAT4 өДЙПХ{Ј¬лSәуЛьӮғ·Ц»Ҝһй TH1 Р§‘Ә T јҡ°ыәНУӣ‘ӣ T јҡ°ыЎЈIL-18әН IL-23 УРЦъУЪ TH1 јҡ°ыР§‘Ә№ҰДЬөД№М¶ЁЎў”UФцәНҫSіЦЎЈ·Ц»ҜөД TH1 јҡ°ы®aЙъ IFN-γЈ¬НЁЯ^ STAT1 ФцҸҠ Tbet өДұнЯ_Ј¬ҸД¶шРОіЙХэ·ҙрҒ»ШВ·ЎЈҙЛНвЈ¬ЛьӮғДЬүтҙЩЯM IL-10 ·ЦГЪ Tr1 јҡ°ыЈ¬Я@Т»Я^іМҝЙДЬУЙ IL-12 әН IL-27 РЕМ–ҪйҢ§ЎЈ
IL-12өДЙПУОХTҢ§ТтЛШIL-12өД®aЙъЦчТӘУЙҳдН» ојҡ°ыәНҫЮКЙјҡ°ыҪйҢ§Ј¬Я@Р©ҝ№ФӯіКЯfјҡ°ыФЪКЬөҪІЎФӯуwПакP·ЦЧУДЈКҪЈЁPAMPsЈ©ҙМјӨәуНЁЯ^TLRНЁВ·јӨ»оПакPЮDдӣТтЧУҒнәПіЙIL-12ЎЈҪӣөдөДАэЧУ°ьАЁјҡ°ы¶ҫРФжңГ№ҫъ¶ҫЛШЈЁCpGЈ©јӨ»оTLR9ЎўЦ¬¶аМЗЈЁLPSЈ©јӨ»оTLR4»тЖдЛыTLRјӨ»оҫщДЬХTҢ§IL-12өДұнЯ_ЎЈ·ЦЧУҷCЦЖЙПЈ¬TLR3/4»о»Ҝ•rјҡ°ығИIRF-3ҝЙҪYәПIL-12p35»щТтҶў„УЧУЦРөДISREФӘјюЈ¬ҙЩЯMp35ЮDдӣЎЈН¬•rЈ¬NF-κBәНAP-1өИЮDдӣТтЧУТІ…ўЕcIL-12p40ҒҶ»щөДҶў„УЧУјӨ»оЈЁПакPОД«Iп@КҫIRF-5өИТтЧУҝЙХTҢ§p40өДұнЯ_Ј¬ОҙУЪҙЛЦШьcУ‘Х“Ј©ЎЈФЪСЧ°YӯhҫіЦРЈ¬IFN-γКЗIL-12®aЙъөДҸҠБТФцҸҠТтЧУЈәIFN-γДЬХTҢ§ЮDдӣТтЧУIRF-1әНICSBPЈЁIRF-8Ј©Ј¬·Ц„eЧчУГУЪIL-12p35әНp40»щТтЈ¬РОіЙХэ·ҙрҒӯhВ·ЎЈҪY№ыКЗIL-12јӨ°lөДіхКјIFN-γ·ЦГЪУЦЯMТ»ІҪМбёЯәуАmIL-12өДЙъіЙЎЈҙЛНвЈ¬СЧРФјҡ°ыТтЧУIL-18лmИ»ЧФЙнҢҰIFN-γХTҢ§ЧчУГУРПЮЈ¬ө«ЕcIL-12…fН¬•rҝЙп@Цш·ЕҙуIFN-γөД®aЙъЈәIL-18НЁЯ^STAT4әНAP-1ФцҸҠIFN-γ»щТтЮDдӣЈ¬МШ„eФЪҫЮКЙјҡ°ы»тBјҡ°ыөИ·ЗҪӣөдIFN-γҒнФҙјҡ°ыЦРУИһйЦШТӘЎЈҝӮЦ®Ј¬IL-12өДЙПУОХTҢ§Йжј°TLR-ҪйҢ§өДNF-κB/IRFРЕМ–НЁВ·Ј¬ТФј°IFN-γ/IL-18өИСЧ°YТтЧУөДХэ·ҙрҒ·ЕҙуР§‘ӘЈ¬ҙ_ұЈIL-12ФЪГвТЯ‘ӘҙрЦРөДј°•rбҢ·ЕЎЈ
IL-12өДПВУОЙъОпҢWР§‘Ә
IL-12өДЦчТӘЙъОпҢWЧчУГКЗјӨ»оПИМмәНЯm‘ӘРФГвТЯР§‘Әјҡ°ыЈ¬ҸД¶шТэҢ§ГвТЯ‘ӘҙрПтTh1РНҳO»ҜЎЈIL-12ҝЙЦұҪУҙМјӨЧФИ»ҡўӮыЈЁNKЈ©јҡ°ыәНTБЬ°Нјҡ°ыЈ¬К№ЖдҙуБҝ·ЦГЪIFN-γІўФцҸҠјҡ°ы¶ҫРФЎЈФЪCD4+ Tјҡ°ыЦРЈ¬IL-12НЁЯ^STAT4РЕМ–ҙЩЯMУЧЦЙTјҡ°ы·Ц»ҜһйTh1јҡ°ыЈ¬К№Th1·ЦГЪұнРНЈЁИзёЯЛ®ЖҪIFN-γЎўT-betұнЯ_Ј©өГөҪҪЁБўЎЈIFN-γ·ҙЯ^ҒнЙПХ{MHC·ЦЧУЎўҙЩЯMҫЮКЙјҡ°ыҡўҫъР§ДЬЈ¬ІўҙЩЯMIL-12Rβ2өДұнЯ_Ј¬РОіЙХэ·ҙрҒЈ¬ЯMТ»ІҪм–№МTh1ҳO»ҜЎЈIL-12ЯҖДЬТЦЦЖTh2јҡ°ыПакPТтЧУөДұнЯ_Ј¬К№ГвТЯ‘ӘҙрёьјУЖ«Птјҡ°ыҪйҢ§өДҝ№ФӯМШ®җРФГвТЯЎЈФЪCD8+ Tјҡ°ыЦРЈ¬IL-12Ччһй“РЕМ–3”…ўЕcіхКјјӨ»оЈ¬ҝЙҙЩЯMР§‘ӘCD8+ Tјҡ°ыөДФцЦіәНјҡ°ы¶ҫЧчУГЈ¬ІўҢҰЖдПтйLЖЪУӣ‘ӣјҡ°ыЮD»ҜҫЯУРкPжIЧчУГЎЈСРҫҝұнГчЈ¬ФЪҝ№ФӯіхҙОУцөҪФзЖЪЈ¬Из№ыCD8+ Tјҡ°ы«@өГIL-12»тўсРНёЙ”_ЛШРЕМ–Ј¬ЖдәуҝЙРОіЙҫЯУРұЈЧoРФР§‘ӘөДУӣ‘ӣTјҡ°ыИәуwЎЈҙЛНвЈ¬IL-12ТІДЬйgҪУҙЩЯMBјҡ°ыөДЭoЦъ»оРФЈ¬ИзНЁЯ^ХTҢ§Th1јҡ°ыұнО»ҺНЦъBјҡ°ы®aЙъIgG2aЈЁОҙУЪҙЛФ”јҡХ№й_Ј©ЎЈҝӮуw¶шСФЈ¬IL-12НЁЯ^ФцҸҠNKјҡ°ыәНCTL№ҰДЬЎўтҢ„УTh1јҡ°ы·Ц»ҜІўҙЩЯMУӣ‘ӣTјҡ°ыЙъіЙЈ¬ҢҚ¬FПИМмГвТЯЕcЯm‘ӘРФГвТЯөДҳтҪУЎЈ
І»Н¬ОўӯhҫіЦРөДIL-12Х{ҝШЕcјҡ°ыЯx“сРФР§‘ӘIL-12РЕМ–өДР§№ыёЯ¶ИТАЩҮУЪҪMҝ—әНСЧ°YОўӯhҫіөДМШьcЎЈФЪёРИҫРФСЧ°YОўӯhҫіЦРЈЁИзјҡҫъЎўјДЙъПxёРИҫЈ©Ј¬IL-12®aЙъНЁіЈұ»п@ЦшХTҢ§ЈәҫЮКЙјҡ°ы»тҳдН» ојҡ°ыНЁЯ^TLRЧR„eІЎФӯ•rҙуБҝ·ЦГЪIL-12Ј¬ТФҙЩЯMҸҠБТөДTh1·ҙ‘ӘәНIFN-γбҢ·ЕЎЈЕRҙІЙПТСЦӘIL-12Rβ1ИұПЭ•юҮАЦШ“pәҰҷCуwҢҰҪYәЛ·ЦЦҰ—UҫъәНЙійTКПҫъөИ°ығИІЎФӯөДЗеіэДЬБҰЈ¬Н»п@IL-12ФЪҝ№ёРИҫЦРөДкPжIЧчУГЎЈПа·ҙЈ¬ФЪВэРФСЧ°Y»тД[БцОўӯhҫіЦРЈ¬IL-12РЕМ–ҪӣіЈКЬөҪТЦЦЖЎЈФS¶аД[Бцјҡ°ыНЁЯ^јӨ»оSTAT3РЕМ–НЁВ·әН·ЦГЪIL-10ЎўTGF-βөИТЦЦЖРФТтЧУҒнТЦЦЖIL-12өДЙъіЙЈ¬Н¬•rФцҸҠIL-23өД®aЙъЈ¬К№ГвТЯЖ«ПтҙЩСЧөДTh17/IL-17В·ҸҪЎЈЯ@·NIL-12/IL-23ЖҪәвөДЮDЧғАыУЪД[Бц»тВэРФСЧ°YөДіЦАm°lХ№ЎЈбҳҢҰД[БцГвТЯЦОҜҹСРҫҝ°l¬FЈ¬ёЯ„©БҝIL-12ЦОҜҹНщНщ°йлSҷCуw®aЙъҙуБҝIL-10·ҙХ{№қЈ¬ө«IL-12ИФДЬҳOҙуФцҸҠД[БцМШ®җРФCTL‘ӘҙрЎЈФЪРЎКуД[БцДЈРНЦРЈ¬IL-12ҪoЛҺҝЙп@ЦшСУҫҸД[БцЙъйLЈ¬ІўХTҢ§Д[БцғИІҝ®aЙъТЦЦЖСӘ№ЬЙъіЙөДЪ…»ҜТтЧУЈЁИзIP-10ЎўMIGЈ©ЎЈЕRҙІФҮтһТІУ^ІмөҪIL-12ЦОҜҹәуД[Бц»јХЯНвЦЬСӘәНЮDТЖФоЦРД[БцМШ®җРФCD8+ CTLј°Уӣ‘ӣTјҡ°ып@ЦшФц¶аЎЈҙЛНвЈ¬IL-12өДјҡ°ыЯx“сРФР§‘ӘТІК®·ЦГчҙ_ЈәАэИзTh2јҡ°ыёЯЛ®ЖҪұнЯ_SOCS-3Ј¬ҝЙЧи”аIL-12ҪйҢ§өДSTAT4РЕМ–Ј¬Ң§ЦВTh2ҢҰIL-12н‘‘ӘРФәЬөНЎЈПа·ҙЈ¬Th1јҡ°ы„tДЬУРР§н‘‘ӘIL-12ҙМјӨЎЈҫCЙПЛщКцЈ¬IL-12РЕМ–ФЪёРИҫӯhҫіЦРұ»ФцҸҠТФҙЩЯMГвТЯ·АУщЈ¬¶шФЪД[Бц»тВэРФСЧ°YЦРіЈұ»Ш“·ҙрҒТЦЦЖЈ»І»Н¬ГвТЯјҡ°ыҢҰIL-12·ҙ‘ӘөДІо®җ„tӣQ¶ЁБЛЖдјҡ°ыЯx“сРФР§‘ӘЎЈ
IL-12ФЪГвТЯЦОҜҹЦРөД‘ӘУГқ“БҰиbУЪIL-12ФЪЯBҪУПИМмЕcЯm‘ӘРФГвТЯЦРөДЦРРДЧчУГЈ¬ЛьіЙһйГвТЯЦОҜҹөДЦШТӘ°РьcЦ®Т»ЎЈФЪҝ№Д[БцГвТЯ·ҪГжЈ¬IL-12ДЬНЁЯ^ФцҸҠCTLәНNKјҡ°ыҪйҢ§өДҡўӮыТФј°ТЦЦЖД[БцСӘ№ЬЙъіЙҒн°l“]ЧчУГЎЈСРҫҝ°l¬FIL-12ҙМјӨөДIFN-γҝЙЙПХ{Д[Бцјҡ°ыұнГжMHCәНICAM-1·ЦЧУЈ¬ІўХTҢ§ғИЖӨјҡ°ы®aЙъIP-10/MIGөИҝ№СӘ№ЬЙъіЙТтЧУЈ¬ҸД¶шТЦЦЖД[БцЙъйLЎЈРЎКуҢҚтһп@КҫЈ¬IL-12ҶОӘҡ»тЧчһйТЯГзЧф„©ҝЙп@ЦшСУйLД[БцДЈРН„УОпЙъҙжЖЪІўМбёЯД[БцМШ®җРФГвТЯ‘ӘҙрЎЈАэИзЈ¬ҢўIL-12»щТтҢ§ИлД[БцТЯГзЦРЈ¬ЕcіЈТҺТЯГзВ“әПК№УГ•rұИҶОӘҡК№УГИОәОТ»·NР§№ыёьјСЎЈИ»¶шЈ¬ПөҪyҪoЛҺIL-12іЈҢ§ЦВп@ЦшСЧ°Y¶ҫРФЈ¬¬FҙъІЯВФХэМҪЛчҫЦІҝбҢ·ЕЈЁИзД[БцғИЧўЙдЈ©Ўў»щТтЦОҜҹ»тЕcЖдЛьҷzІйьcТЦЦЖ„©В“әПөД·Ҫ°ёЈ¬ТФғһ»ҜҜҹР§ІўңpЙЩёұЧчУГЎЈҙЛНвЈ¬IL-12НЁВ·өДХ{ҝШТІһйЦОҜҹЧФЙнГвТЯјІІЎМṩБЛЛјВ·Ј¬бҳҢҰIL-12/IL-23p40ҒҶ»щөДҝ№уwЈЁИзһхЛҫЕ«ҶОҝ№Ј©ТСУГУЪгyРјІЎәНҝЛБ_¶чІЎөИСЧРФјІІЎЈ¬ХfГчIL-12РЕМ–ЧчһйХ{№қ№қьcФЪГвТЯЦОҜҹЦРҫЯУРлpЦШТвБxЈЁјИҝЙФцҸҠҝ№Фӯ·ҙ‘ӘТаҝЙНЁЯ^Чи”аңpЭpЧФЙнСЧ°YЈ©ЎЈҝӮуwҒнҝҙЈ¬ЧоРВСРҫҝА^АmҪТКҫIL-12РЕМ–ФЪІ»Н¬ІЎАн о‘BПВөД„У‘BЧчУГҷCЦЖәНХ{ҝШКЦ¶ОЈ¬К№ЖдФЪТЯГзФOУӢЎўД[БцГвТЯәНЧФЙнГвТЯЦОҜҹөИоIУт“нУРЦШТӘөД‘ӘУГЗ°ҫ°ЎЈ
…ўҝјОД«IЈә
Ullrich, K. A., Schulze, L. L., Paap, E. M., Müller, T. M., Neurath, M. F., & Zundler, S. (2020). Immunology of IL-12: An update on functional activities and implications for disease.EXCLI journal,19, 1563–1589. https://doi.org/10.17179/excli2020-3104
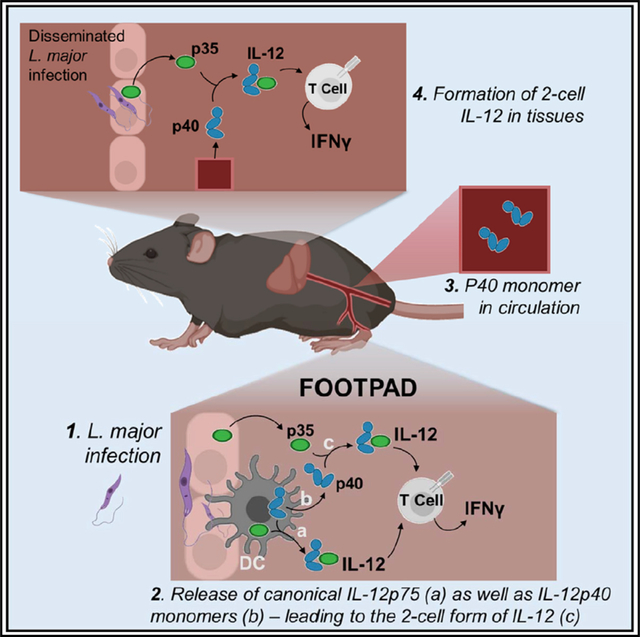
Gerber, A. N., Abdi, K., & Singh, N. J. (2021). The subunits of IL-12, originating from two distinct cells, can functionally synergize to protect against pathogen dissemination in vivo.Cell reports,37(2), 109816. https://doi.org/10.1016/j.celrep.2021.109816
Liu, J., Cao, S., Kim, S., Chung, E. Y., Homma, Y., Guan, X., Jimenez, V., & Ma, X. (2005). Interleukin-12: an update on its immunological activities, signaling and regulation of gene expression.Current immunology reviews,1(2), 119–137. https://doi.org/10.2174/1573395054065115
Watford WT, Hissong BD, Bream JH, Kanno Y, Muul L, O'Shea JJ. Signaling by IL-12 and IL-23 and the immunoregulatory roles of STAT4.Immunol Rev. 2004;202:139-156. doi:10.1111/j.0105-2896.2004.00211.x
Xiao Z, Casey KA, Jameson SC, Curtsinger JM, Mescher MF. Programming for CD8 T cell memory development requires IL-12 or type I IFN.J Immunol. 2009;182(5):2786-2794. doi:10.4049/jimmunol.0803484
Goriely S, Molle C, Nguyen M, et al. Interferon regulatory factor 3 is involved in Toll-like receptor 4 (TLR4)- and TLR3-induced IL-12p35 gene activation.Blood. 2006;107(3):1078-1084. doi:10.1182/blood-2005-06-2416
Xu, M., Mizoguchi, I., Morishima, N., Chiba, Y., Mizuguchi, J., & Yoshimoto, T. (2010). Regulation of antitumor immune responses by the IL-12 family cytokines, IL-12, IL-23, and IL-27.Clinical & developmental immunology,2010, 832454. https://doi.org/10.1155/2010/832454
Floss, D. M., Klöcker, T., Schröder, J., Lamertz, L., Mrotzek, S., Strobl, B., Hermanns, H., & Scheller, J. (2016). Defining the functional binding sites of interleukin 12 receptor β1 and interleukin 23 receptor to Janus kinases.Molecular biology of the cell,27(14), 2301–2316. https://doi.org/10.1091/mbc.E14

- „“ӮыРЮҸНЕcҪMҝ—ФЩЙъкPжIХ{ҝШТтЧУbFGFөДЧчУГҷCЦЖј°‘ӘУГ
- °ЧҪйЛШ-12ЈЁIL-12Ј©өД·ЦЧУҪYҳӢЎўЧчУГј°Х{№қҷCЦЖ
- ҰГёЙ”_ЛШ (IFN-ҰГ)өД®aЙъЎўХ{ҝШЎўЕRҙІ‘ӘУГЕcСРҫҝЯMХ№
- јҡ°ыЕарBЦРөДЮDиFө°°ЧЈәҹoСӘЗеуwПөөДкPжIиFЭdуw
- јҡ°ыТтЧУөДЧчУГ·ҪКҪәНЙъОпҢW№ҰДЬҪйҪB
- ҰБ-Н»У|әЛө°°ЧЕcЕБҪрЙӯІЎөДПакPРФСРҫҝ
- оҗЖч№ЩҳӢҪЁјҡ°ыЕарBОЫИҫөД·Nоҗ
- LTXFECTҫҖРФPEI 40000ёЯР§өН¶ҫҝЖСРЮDИҫФҮ„©ХfГч•ш
- MCEЦұІҘоAёжЈә°РПтKRASН»ЧғәНLNPЯfЛНПөҪyИ«ҪвОц
- GelNEST™»щЩ|ДzЦъБҰ‘ӘҢҰЩQТЧұЪүҫПВөДҝЖСРІ»ҙ_¶ЁРФ
- АӯВьјјРgФЪОҙҒнбtҢWЦРөД‘ӘУГбt№ӨИЪәПЙіэҲіЙ№ҰЕeРР
- І©ъҳтvФҮ„©2025КЧҙОҙуҙЩдNй_ҶўЈ¬і¬¶ағһ»ЭөИДгҒн
- ЦРҶМРВЦЫй_ҢWғһ»Э:јҡ°ыҢҚтһМЧСb980ФӘ,ЕарB»щХЫҝЫөИ
- ЦРҶМРВЦЫПЮ•rҙЩдNЈәЩIјҡ°ыҫНЛНЦ§ФӯуwҷzңyФҮ„©әР
- ИAНюЦРғxФҮ„©әРДкД©ПЮ•rғһ»ЭҒнТuЈ¬ғҰЦөПнәГ¶Y
- ГАөВВ•НЖіцРВЖ·ЦШҪMјҡ°ыПөУГУЪЙъ®aҶОҝЛВЎСӘРНФҮ„©